Why can you possess traits neither of your parents have? The relationship of genotype to phenotype is rarely as simple as the dominant and recessive patterns described by Mendel.
Aa Aa Aa
One of Gregor Mendel's greatest contributions to the study of heredity was the concept of dominance. Mendel observed that a heterozygote offspring can show the same phenotype as the parent homozygote, so he concluded that there were some traits that dominated over other inherited traits. However, the relationship of genotype to phenotype is rarely as simple as the dominant and recessive patterns described by Mendel. As the study of inheritance expanded beyond the seven traits Mendel initially examined and also included organisms other than pea plants, biologists began to notice a variety of relationships between alleles that code for the same trait. These allelic interactions were not exclusively recessive or dominant, and they greatly enriched our understanding of how genotype leads to phenotype.
Complete versus Partial Dominance
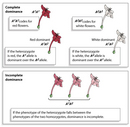
 Figure 1 Figure Detail
Figure 1 Figure Detail
Dominance affects the phenotype derived from an organism's genes, but it does not affect the way these genes are inherited. Complete dominance occurs when the heterozygote phenotype is indistinguishable from that of the homozygous parent. However, sometimes the heterozygote displays a phenotype that is an intermediate between the phenotypes of both homozygote parents (one of which is homozygous dominant, and the other of which is homozygous recessive). This intermediate phenotype is a demonstration of partial or incomplete dominance. When partial dominance occurs, a range of phenotypes is usually observed among the offspring. Although the offspring may show a variety of phenotypes, each one will lie along a continuum bracketed by the homozygous parental phenotypes.
In Figure 1, for example, neither flower color (red or white) is fully dominant. Thus, when homozygous red flowers (A1A1) are crossed with homozygous white (A2A2), a variety of pink-shaded phenotypes result. Note, however, that partial dominance is not the same as blending inheritance; after all, when two F1 pink flowers are crossed, both red and white flowers are found among the progeny. In other words, nothing is different about the way these alleles are inherited; the only difference is in the way the alleles determine phenotype when they are combined.
Codominance
As opposed to partial dominance, codominance occurs when the phenotypes of both parents are simultaneously expressed in the same offspring organism. Indeed, "codominance" is the specific term for a system in which an allele from each homozygote parent combines in the offspring, and the offspring simultaneously demonstrates both phenotypes. An example of codominance occurs in the human ABO blood group system. Many blood proteins contribute to blood type (Stratton, 1952), and the ABO protein system in particular defines which types of blood you can receive in a transfusion. In a hospital setting, attention to the blood proteins present in an individual's blood cells can make the difference between improving health and causing severe illness.
There are three common alleles in the ABO system. These alleles segregate and assort into six genotypes, as shown in Table 1.
Table 1: Possible ABO Genotypes
Blood Type Related Genotype(s) A AA or AO B BB or BO AB AB O OO As Table 1 indicates, only four phenotypes result from the six possible ABO genotypes. How does this happen? To understand why this occurs, first note that the A and B alleles code for proteins that exist on the surface of red blood cells; in contrast, the third allele, O, codes for no protein. Thus, if one parent is homozygous for type A blood and the other is homozygous for type B, the offspring will have a new phenotype, type AB. In people with type AB blood, both A and B proteins are expressed on the surface of red blood cells equally. Therefore, this AB phenotype is not an intermediate of the two parental phenotypes, but rather is an entirely new phenotype that results from codominance of the A and B alleles. Of course, in this system, AB is not the only heterozygote; people may also be AO or BO. All of these heterozygote genotypes demonstrate the coexistence of two phenotypes within the same individual.
Overdominance
In some instances, offspring can demonstrate a phenotype that is outside the range defined by both parents. In particular, the phenomenon known as overdominance occurs when a heterozygote has a more extreme phenotype than that of either of its parents. Indeed, in a few examples, a trait that shows overdominance sometimes confers a survival advantage in the heterozygote (Parsons & Bodmer, 1961).
A well-known example of overdominance occurs in the alleles that code for sickle-cell anemia. Sickle-cell anemia is a debilitating disease of the red blood cells, wherein a single amino acid deletion causes a change in the conformation of a person's hemoglobin such that the person's red blood cells are elongated and somewhat curved, taking on a sickle shape. This change in shape makes the sickle red blood cells less efficient at transporting oxygen through the bloodstream.
The altered form of hemoglobin that causes sickle-cell anemia is inherited as a codominant trait. Specifically, heterozygous (Ss) individuals express both normal and sickle hemoglobin, so they have a mixture of normal and sickle red blood cells. In most situations, individuals who are heterozygous for sickle-cell anemia are phenotypically normal. Under these circumstances, sickle-cell disease is a recessive trait. Individuals who are homozygous for the sickle-cell allele (ss), however, may have sickling crises that require hospitalization. In severe cases, this condition can be lethal.
Producing altered hemoglobin can be beneficial for inhabitants of countries afflicted with falciparum malaria, an extremely deadly parasitic disease. Sickle blood cells "collapse" around the parasites and filter them out of the blood. Thus, people who carry the sickle-cell allele are more likely to recover from malarial infection. In terms of combating malaria, the Ss genotype has an advantage over both the SS genotype, because it results in malarial resistance, and the ss genotype, because it does not cause sickling crises. This complex example of overdominance may be the sole reason that the allele persists in the human population today (Keeton & Gould, 1986).
Multiple Alleles and Dominance Series
Allelic dominance always depends on the relative influence of each allele for a specific phenotype under certain environmental conditions. For example, in the pea plant (Pisum sativum), the timing of flowering follows a monohybrid (single-gene) inheritance pattern in certain genetic backgrounds. While there is some variation in the exact time of flowering within plants that have the same genotype, specific alleles at this locus (Lf) can exert temporal control of flowering in different backgrounds (Murfet, 1975). Investigators have found evidence for four different alleles at this locus: Lfd, Lf, lf,and lfa. Plants homozygous for the lfa allele flower the earliest, while Lfd plants flower the latest. A single allele causes the delayed flowering. Thus, the multiple alleles at the Lf locus represent an allelic series, with each allele being dominant over the next allele in the series. In this case: Lfd > Lf > lf > lfa (Table 2).
Table 2: Flowering Times in Pisum sativum with Different Lf Alleles
Genotype Earliest Flowering Early Intermediate Flowering Late Intermediate Flowering Latest Flowering Lf Lf + Lfd Lfd + Lfd Lf + lfa lfa + lf lf + lf lfa + Lf lfa +
Summarizing the Role of Dominance and Recessivity
Mendel's early work with pea plants provided the foundational knowledge for genetics, but Mendel's simple example of two alleles, one dominant and one recessive, for a given gene is a rarity. In fact, dominance and recessiveness are not actually allelic properties. Rather, they are effects that can only be measured in relation to the effects of other alleles at the same locus. Furthermore, dominance may change according to the level of organization of the phenotype. Variations of dominance highlight the complexity of understanding genetic influences on phenotypes.
Aa Aa Aa
One of Gregor Mendel's greatest contributions to the study of heredity was the concept of dominance. Mendel observed that a heterozygote offspring can show the same phenotype as the parent homozygote, so he concluded that there were some traits that dominated over other inherited traits. However, the relationship of genotype to phenotype is rarely as simple as the dominant and recessive patterns described by Mendel. As the study of inheritance expanded beyond the seven traits Mendel initially examined and also included organisms other than pea plants, biologists began to notice a variety of relationships between alleles that code for the same trait. These allelic interactions were not exclusively recessive or dominant, and they greatly enriched our understanding of how genotype leads to phenotype.
Complete versus Partial Dominance
Dominance affects the phenotype derived from an organism's genes, but it does not affect the way these genes are inherited. Complete dominance occurs when the heterozygote phenotype is indistinguishable from that of the homozygous parent. However, sometimes the heterozygote displays a phenotype that is an intermediate between the phenotypes of both homozygote parents (one of which is homozygous dominant, and the other of which is homozygous recessive). This intermediate phenotype is a demonstration of partial or incomplete dominance. When partial dominance occurs, a range of phenotypes is usually observed among the offspring. Although the offspring may show a variety of phenotypes, each one will lie along a continuum bracketed by the homozygous parental phenotypes.
In Figure 1, for example, neither flower color (red or white) is fully dominant. Thus, when homozygous red flowers (A1A1) are crossed with homozygous white (A2A2), a variety of pink-shaded phenotypes result. Note, however, that partial dominance is not the same as blending inheritance; after all, when two F1 pink flowers are crossed, both red and white flowers are found among the progeny. In other words, nothing is different about the way these alleles are inherited; the only difference is in the way the alleles determine phenotype when they are combined.
Codominance
As opposed to partial dominance, codominance occurs when the phenotypes of both parents are simultaneously expressed in the same offspring organism. Indeed, "codominance" is the specific term for a system in which an allele from each homozygote parent combines in the offspring, and the offspring simultaneously demonstrates both phenotypes. An example of codominance occurs in the human ABO blood group system. Many blood proteins contribute to blood type (Stratton, 1952), and the ABO protein system in particular defines which types of blood you can receive in a transfusion. In a hospital setting, attention to the blood proteins present in an individual's blood cells can make the difference between improving health and causing severe illness.
There are three common alleles in the ABO system. These alleles segregate and assort into six genotypes, as shown in Table 1.
Table 1: Possible ABO Genotypes
Blood Type Related Genotype(s) A AA or AO B BB or BO AB AB O OO As Table 1 indicates, only four phenotypes result from the six possible ABO genotypes. How does this happen? To understand why this occurs, first note that the A and B alleles code for proteins that exist on the surface of red blood cells; in contrast, the third allele, O, codes for no protein. Thus, if one parent is homozygous for type A blood and the other is homozygous for type B, the offspring will have a new phenotype, type AB. In people with type AB blood, both A and B proteins are expressed on the surface of red blood cells equally. Therefore, this AB phenotype is not an intermediate of the two parental phenotypes, but rather is an entirely new phenotype that results from codominance of the A and B alleles. Of course, in this system, AB is not the only heterozygote; people may also be AO or BO. All of these heterozygote genotypes demonstrate the coexistence of two phenotypes within the same individual.
Overdominance
In some instances, offspring can demonstrate a phenotype that is outside the range defined by both parents. In particular, the phenomenon known as overdominance occurs when a heterozygote has a more extreme phenotype than that of either of its parents. Indeed, in a few examples, a trait that shows overdominance sometimes confers a survival advantage in the heterozygote (Parsons & Bodmer, 1961).
A well-known example of overdominance occurs in the alleles that code for sickle-cell anemia. Sickle-cell anemia is a debilitating disease of the red blood cells, wherein a single amino acid deletion causes a change in the conformation of a person's hemoglobin such that the person's red blood cells are elongated and somewhat curved, taking on a sickle shape. This change in shape makes the sickle red blood cells less efficient at transporting oxygen through the bloodstream.
The altered form of hemoglobin that causes sickle-cell anemia is inherited as a codominant trait. Specifically, heterozygous (Ss) individuals express both normal and sickle hemoglobin, so they have a mixture of normal and sickle red blood cells. In most situations, individuals who are heterozygous for sickle-cell anemia are phenotypically normal. Under these circumstances, sickle-cell disease is a recessive trait. Individuals who are homozygous for the sickle-cell allele (ss), however, may have sickling crises that require hospitalization. In severe cases, this condition can be lethal.
Producing altered hemoglobin can be beneficial for inhabitants of countries afflicted with falciparum malaria, an extremely deadly parasitic disease. Sickle blood cells "collapse" around the parasites and filter them out of the blood. Thus, people who carry the sickle-cell allele are more likely to recover from malarial infection. In terms of combating malaria, the Ss genotype has an advantage over both the SS genotype, because it results in malarial resistance, and the ss genotype, because it does not cause sickling crises. This complex example of overdominance may be the sole reason that the allele persists in the human population today (Keeton & Gould, 1986).
Multiple Alleles and Dominance Series
Allelic dominance always depends on the relative influence of each allele for a specific phenotype under certain environmental conditions. For example, in the pea plant (Pisum sativum), the timing of flowering follows a monohybrid (single-gene) inheritance pattern in certain genetic backgrounds. While there is some variation in the exact time of flowering within plants that have the same genotype, specific alleles at this locus (Lf) can exert temporal control of flowering in different backgrounds (Murfet, 1975). Investigators have found evidence for four different alleles at this locus: Lfd, Lf, lf,and lfa. Plants homozygous for the lfa allele flower the earliest, while Lfd plants flower the latest. A single allele causes the delayed flowering. Thus, the multiple alleles at the Lf locus represent an allelic series, with each allele being dominant over the next allele in the series. In this case: Lfd > Lf > lf > lfa (Table 2).
Table 2: Flowering Times in Pisum sativum with Different Lf Alleles
Genotype Earliest Flowering Early Intermediate Flowering Late Intermediate Flowering Latest Flowering Lf Lf + Lfd Lfd + Lfd Lf + lfa lfa + lf lf + lf lfa + Lf lfa +
Summarizing the Role of Dominance and Recessivity
Mendel's early work with pea plants provided the foundational knowledge for genetics, but Mendel's simple example of two alleles, one dominant and one recessive, for a given gene is a rarity. In fact, dominance and recessiveness are not actually allelic properties. Rather, they are effects that can only be measured in relation to the effects of other alleles at the same locus. Furthermore, dominance may change according to the level of organization of the phenotype. Variations of dominance highlight the complexity of understanding genetic influences on phenotypes.

")